Book review: “The Heart of the Brain: The Hypothalamus and Its Hormones”
1. Introduction
1.1 Hypothalamus as “business logic”
In software jargon, there’s a nice term “business logic”, for code like the following (made-up) excerpt from corporate tax filing software (based on here):
def attachSupplementalDocuments(file):
if file.state == "California" or file.state == "Texas":
# SR008-04X/I are always required in these states
file.attachDocument("SR008-04X")
file.attachDocument("SR008-04XI")
if file.ledgerAmnt >= 500_000:
# Ledger of 500K or more requires AUTHLDG-1A
file.attachDocument("AUTHLDG-1A")When you think of “business logic”, think of stuff like that—i.e., parts of source code that more-or-less directly implement specific, real-world, functional requirements.
By contrast, things that are NOT business logic include infrastructure & subroutines & plumbing that are generally useful in many contexts—e.g. code for initializing a database, or code for memory management, or code for performing stochastic gradient descent.
If genomes are the “source code” of brains, then they need to encode “business logic” too—specific calculations to do specific things that help an animal thrive and reproduce in its particular biological niche. For example:
if about_to_starve_to_death:
reduce_sex_drive()
increase_pain_tolerance()
emit_hunger_sensation()
increase_reward_for_eating()
if fertility > 0:
increase_sex_drive()
...(We could also call these things “innate reactions”.)
Machine Learning people might interject here: Why does that have to be in the genome? Why can’t the brain derive those kinds of rules via a within-lifetime learning algorithm instead? Well, often it does! But:
Some things can’t be learned within a lifetime, because their evolutionary benefits are only apparent in hindsight, perhaps even many generations hence.
Other things can’t be learned within a lifetime, because not doing them, even once, is potentially fatal.
Still other bits of “business logic” involves setting up the animal’s within-lifetime learning algorithms. A particularly important example in this category is that animals learn (in part) by reinforcement learning, and reinforcement learning needs a reward function—the thing that says eating yummy food is good and touching a hot stove is bad, as opposed to the other way around. Calculating that reward function involves a lot of “business logic”.
Anyway, the genome puts much of the brain’s “business logic” into a part of the brain called the hypothalamus, the subject of this post. (I’ll walk through an example in Section 3 below.) The “business logic” doesn’t all go into the hypothalamus—the brainstem gets some too.[1] But I believe that, apart from the hypothalamus and brainstem, the other 96% (!) of human brain volume has essentially no “business logic” at all, but rather is dedicated to running within-lifetime learning algorithms—see my earlier post “Learning from scratch” in the brain.
In his book, Gareth Leng poetically gives some examples of what the hypothalamus does for us:
When we are dehydrated, [the hypothalamus] makes us thirsty and tells our kidney not to waste water but to concentrate our urine. When we are ill, it raises our body temperature, generating a fever that kills off infections. When our blood sugar is low, it tells us to eat, and when our stomach is full, to stop. It determines the shape of our bodies, how tall we will grow, how fat we will be, and where our fat and muscle will grow. When we are frightened or anxious or stressed it determines whether we will set our teeth, stiffen our sinews, freeze, or fight—or flee. The rhythms of days and of seasons are beaten by its drums, and we grow and attain puberty under its tutelage. And, led by the hypothalamus, we preen and woo, and the sap rises in our loins; a man produces sperm, and in a woman the ovarian cycle turns; we court and mate and bond. A woman conceives, and her body changes to the needs of the child within, and she gives birth and produces milk, and she loves and nurtures her children.
I’ll get back to “business logic” via the Case Study in Section 3; first, we have more neuroscience-y background to cover.
1.2 Neuroanatomy
Here’s the hypothalamus:
{kind=link}
As Leng puts it, “If you curl your tongue back as far as you can and press it on the roof of your mouth, the hypothalamus will be almost on the tip of your tongue.”
For all that it does, the human hypothalamus is pretty small—4cm³, or 0.3% of adult human brain volume (ref). Like almost everything in the brain, this total is split into two somewhat-mirror-symmetric copies on the left and right. I can’t find any sources saying how many neurons it has.[2]
1.3 Relation to pituitary gland
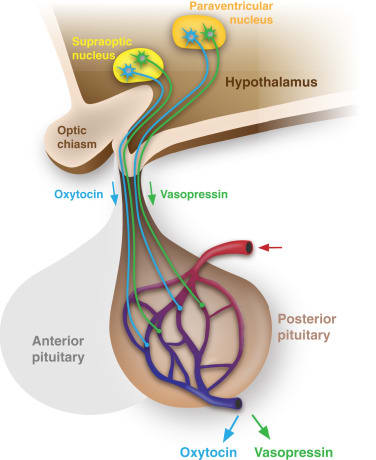
The diagram above also shows the pituitary gland below the hypothalamus. The hypothalamus and pituitary have a very close relationship. As an example, here’s the weirdly-indirect way that the brain releases the stress hormone cortisol:
First, the hypothalamus squirts a tiny amount of CRH (Corticotropin-releasing hormone) into the hypophyseal portal system, a system of tiny blood vessels connecting part of the hypothalamus to the pituitary gland.
Next, in the pituitary gland, the CRH triggers the release of a larger amount of ACTH (adrenocorticotropic hormone) into the general bloodstream
Finally, in the adrenal gland (near the kidneys), the circulating ACTH in turn triggers the release of a still-larger amount of cortisol into the same general bloodstream
(This chain is called the “HPA (hypothalamus-pituitary-adrenal) axis”.)
The hypophyseal portal system (the tiny blood vessels in the first step) comprises a small volume of blood, which allows the puppet-master hypothalamus to control the anterior pituitary by squirting out a truly minuscule quantity of CRH or other “release factor” hormones. Nonetheless, in the 1960s-80s, three teams of scientists (led by Andrew Schally, Roger Guillemin, and Wylie Vale) figured out a technique for identifying these hormones. What’s the trick? Oh, I’m glad you asked. Here’s Schally’s Nobel prize lecture:
Oscar Mayer & Co. generously donated about a million pig hypothalami. This enabled us to undertake a large-scale effort aimed at the purification of adequate amounts of material to permit chemical characterization … The first isolation of 800 µg [GnRH] from ventral hypothalami of 165,000 pigs was achieved by twelve successive purification steps…
😳
Moving on, the adult human pituitary has an anterior part and a posterior part. The “HPA axis” above involves the anterior pituitary, and so do the analogous “hypothalamic–pituitary–gonadal axis” and “hypothalamic–pituitary–thyroid axis”.
So I still need to talk about the posterior pituitary. This is where two all-important hormones, oxytocin and vasopressin (more on which below), are released into the bloodstream. The hormones are produced up in the hypothalamus, by its “magnocellular” neurons, so-called because they are among the largest and most energy-consuming neurons in the brain. No surprise—they’re tasked with producing enough oxytocin and vasopressin to spread through the general circulation and impact the whole body. (For those keeping track, these magnocellular neurons can be found in the hypothalamus’s “supraoptic nucleus”, and its “paraventricular nucleus”, plus a few more neurons scattered between them.) These neurons have axons that go down to the posterior pituitary; the oxytocin or vasopressin travels down the axons, and is then ejected into the bloodstream during neuron spikes.
1.4 Hypothalamus substructure
The obvious substructure of the hypothalamus is the one that you see under a microscope. The textbooks list maybe a dozen major nuclei—including the arcuate, preoptic, supraoptic, suprachiasmatic, lateral, ventromedial, and supramammillary nuclei. Indeed, just to make sure that the med students suffer, the hypothalamus has both a paraventricular nucleus and a periventricular nucleus!
A 2019 study led by the indefatigable Larry Swanson broke down the hypothalamus further into 65 regions (130 between both hemispheres). After combing the literature and filling in the gaps with their own experiments, they conclude that “The dataset indicated the existence of 7,982 (of 16,770 possible) intrahypothalamic macroconnections”, as illustrated by the following map:
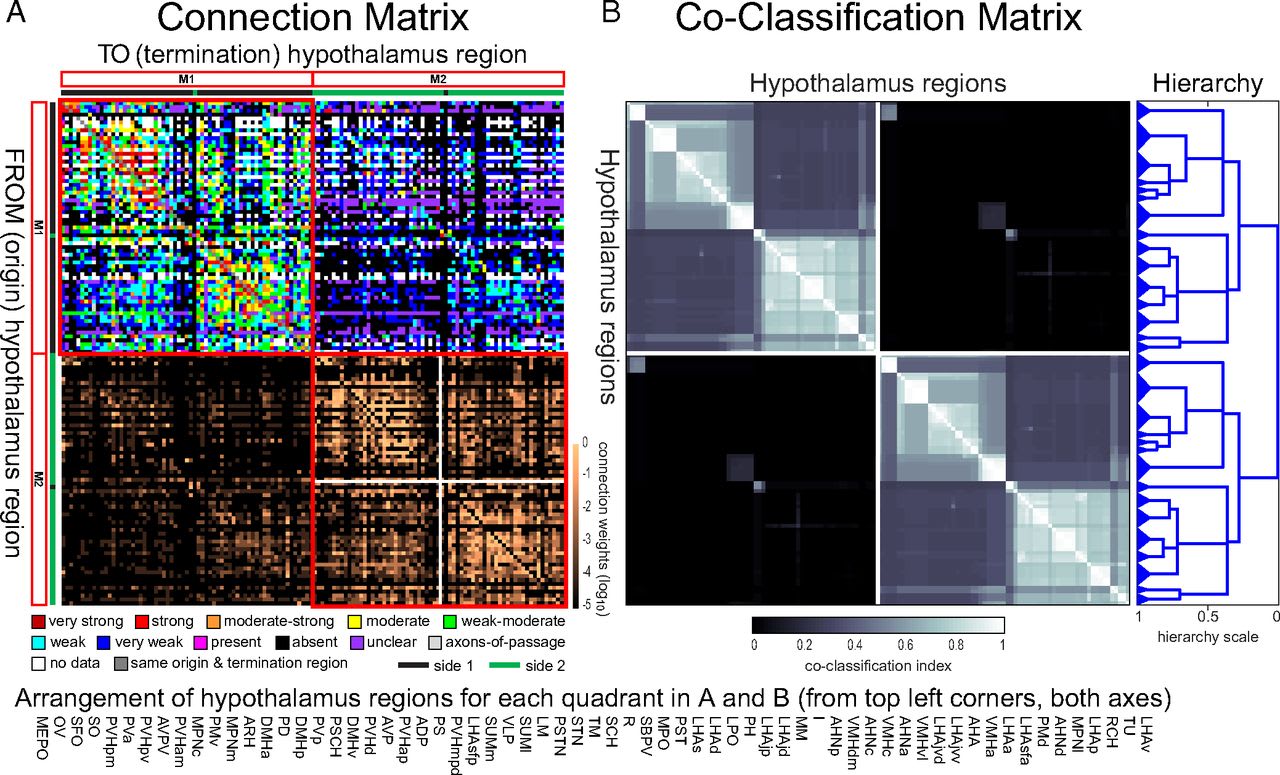
Clear as mud, right?
I initially tried making Anki flashcards for the major hypothalamic nuclei. It worked great in certain cases where the nucleus did just one big thing—for example, the suprachiasmatic nucleus creates circadian rhythm, and the supraoptic nucleus is a factory for oxytocin and vasopressin as mentioned above. In other cases, I was stymied by the fact that one nucleus would perform a laundry list of different functions with no obvious-to-me, memorable pattern. For example, the arcuate nucleus has something to do with feeding, metabolism, fertility, cardiovascular regulation, and much much more. I found it impossible to memorize these lists, and doing so seemed pointless anyway.
I later learned the reason: the most important substructure of the hypothalamus is invisible under the microscope. Instead, it lies in subpopulations of neurons defined not by location but rather by which “neuropeptides” they produce and respond to.
Indeed, it’s hard to say anything about the hypothalamus without talking about neuropeptides. So let’s turn to those next.
2. Neuropeptides
2.1 Introduction to neuropeptides
I already mentioned oxytocin and vasopressin; these are two examples of a much larger class of molecules called “neuropeptides”.
A “peptide” is a short chain of amino acids—basically, a tiny protein. A “neuropeptide” is any peptide that’s used for signaling within the brain.
Here’s Leng with some of the basics on neuropeptides:
A peptide is a chain of amino acids, and which particular amino acids and in which order they are assembled is determined by the gene for that peptide. Oxytocin has nine amino acids and a molecular weight of about 1,000 daltons, and the oxytocin gene must produce it as a fragment of a much larger precursor peptide, which has a molecular weight of about 23,000 daltons. The precursor contains the oxytocin sequence and other sequences that determine what the cell will do with the oxytocin. Part of the precursor determines that it will be packaged into vesicles to be transported to sites where the vesicles can be released. Another part is important for folding the precursor in a way that enables it to aggregate with other precursor molecules so they can be packaged compactly in a vesicle. The vesicles also contain enzymes that cleave oxytocin from the rest of the precursor. It’s complicated and expensive, but the final product is a powerful molecule: oxytocin will survive in the extracellular fluid for much longer than ordinary neurotransmitters, it can act on cells at much lower concentrations, and it acts at sophisticated receptors that have a range of properties through which they control complex signaling pathways within those target cells.…
One striking characteristic of peptides is their ability to orchestrate behavior—to coordinate different systems to evoke a coherent, adaptive, organismal response, be it maternal behavior, aggression, sexual arousal, or behaviors associated with hunger and thirst: foraging, feeding, and drinking. An injection of NPY (neuropeptide Y) into the brain will provoke feeding, and an injection of α-MSH (α-melanocyte-stimulating hormone) will stop it.…
We now know that more than a hundred neuropeptides are expressed in different neuronal populations, and that most if not all neurons in the entire brain release one or more peptide messengers as well as a conventional neurotransmitter. Because peptides have a long half-life and act at receptors at very low concentrations, their actions are not confined to targets adjacent to the site of release. Importantly, peptides in the brain often have organizational and activational roles that seem more like the roles of hormones in the periphery.
The two most famous neuropeptides, oxytocin and vasopressin, differ by two amino acids, and split off from a common ancestor neuropeptide 400 million years ago, back in the good old days when our ancestors were jawless fish. The ancestral form goes back even further, to before our common ancestor with insects—see neurohypophysial hormones. Both oxytocin and vasopressin are involved in social, sexual, and reproductive behaviors, among other things. A handy grossly-oversimplified stereotype that I got from Panksepp is that vasopressin tends to be more involved in male behavior (e.g. intermale aggression) and oxytocin in female behavior (e.g. lactation). But don’t take this too far. For example, in humans, vasopressin is involved in adjusting the composition of urine in the kidneys—an activity which, I am told, is enjoyed equally by both sexes.
2.2 “Clans”
Here’s Leng again:
Neurons of the hypothalamus … comprise many subpopulations—tribes, if you like. Living as I do in Scotland, I’d rather think of clans. Members of a clan are all different from each other, but are more like each other than like members of other clans. Each clan has its characteristic phenotype, dictated by the “tartan” of genes that it wears. Different genes make some neurons sensitive to glucose, temperature, or osmotic pressure, or to particular hormonal signals from the periphery. Others determine the signals that neurons of a clan generate. Different clans can be defined by the signals they use to communicate and the signals to which they can respond: different clans use different combinations of peptides as chemical signals along with “classical” neurotransmitters. These combinations come from more than a hundred known neuropeptides, and to these we should add yet more signaling molecules—prostaglandins, neurosteroids (steroids synthesized in the brain itself), endocannabinoids (endogenous, cannabis-like molecules), and gases like nitric oxide and carbon monoxide. Yet other genes determine where the clan is in the hypothalamus, and the shapes and connectivity of its members.
Some clans regulate the autonomic nervous system, which controls blood pressure, heart rate, digestion, respiration, urination, and sexual arousal. Some others are neuroendocrine neurons: these regulate the secretion of hormones from the pituitary gland—many of which control the secretion of other hormones, including those from the ovary and testes, the adrenal and thyroid glands—and also hormones from the liver, kidneys, gut, and heart. They control not only the functions of organs in our body, but also our behavior, by their actions on other parts of the brain. The hypothalamus of a male is not the same as that of a female—it is a sexually dimorphic structure. It is also plastic—its structure and functions are malleable, and alter according to physiological needs: after puberty, in pregnancy and lactation, in cold and hunger.
Behaviors important to who we are—love and hate, how much we eat and what we eat, how we respond to threat and to stress—are governed by the hypothalamus, and not by the map of how the neurons are connected, but by where the receptors for these peptide signals are found. Neurotransmitter signals are ephemeral and confined by anatomical connectivity, but the peptide signals that hypothalamic neurons generate are potent, wide-reaching, and long-lasting, and they affect not just neuronal signaling but also the genes that neurons express. Remarkably, different peptides when injected into the brain induce coherent, meaningful behaviors—some, for example, trigger eating, others induce a longing for salt or initiate maternal behavior or aggression or sleep.
I mentioned above my failed attempt to make an Anki flashcard for what the arcuate nucleus of the hypothalamus does. Well, the reason is that the arcuate nucleus isn’t one thing, but rather at least a dozen subpopulation of neurons, each doing its own thing with its own particular suite of neuropeptides, as Leng explains:
The arcuate nucleus is at the base of the hypothalamus adjacent to the median eminence. It hosts three populations of neuroendocrine neurons: one secretes GHRH, which controls growth hormone secretion. Another secretes dopamine to regulate prolactin secretion; these fire in synchronous bursts every minute, and also express a peptide, met-enkephalin. Another population of dopamine neurons innervates the intermediate lobe of the pituitary to regulate α-MSH secretion into the blood. Other neurons control feeding: one population makes NPY and AgRP, both of which stimulate feeding, while another makes three peptides: α-MSH,which, in the brain, stimulates sexual behavior and inhibits feeding; the opioid peptide beta-endorphin; and CART (cocaine- and amphetamine-related transcript), which also inhibits feeding. Yet another neuronal population makes three more peptides kisspeptin, neurokinin B, and dynorphin, packaged in three different populations of vesicles. These regulate the pulsatile secretion of GnRH. Yet another makes somatostatin, and there is some evidence that another makes ghrelin. This exuberance is not unusual, and the list is far from exhaustive even for the arcuate nucleus. Most nuclei in the hypothalamus contain many clans of neurons that perform different physiological functions and express a diversity of peptides in addition to classical neurotransmitters.
Is there a unifying theme here, or is it (for all intents and purposes) a meaningless happenstance that these dozen subpopulations all happen to be co-located in the arcuate nucleus, as opposed to somewhere else within the hypothalamus? It’s probably at least partly meaningless. But an exception in this case is that the arcuate nucleus happens to have a nice coastal location on the shores of the hypophyseal portal system (mentioned above), with a correspondingly modified blood-brain barrier. That’s why it houses various neuron groups that need to send signals through the blood to the anterior pituitary gland, along with other neuron groups that need to detect hormones (or other stuff) from the general bloodstream.
In Leng’s “clan” analogy, I guess we’d say that the arcuate nucleus is a town that happens to have a very nice sheltered harbor, so it houses a clan of shipbuilders, a clan of swimmers, a clan of marine biologists, a fishing clan, and so on. So there’s a reason that these clans all find themselves living in the same town, but it’s still fundamentally lots of different clans doing different unrelated things.
2.3 If normal neurotransmitters are “whispered secrets” from one neuron to another, then neuropeptides are “public announcements” to a whole region
Leng returns to this “whispered secrets vs public announcements” maxim throughout the book. One justification is that any given neuron can only release neuropeptides infrequently and in correspondingly large quantities. Here’s Leng with the details:
At a classical synapse, each spike typically releases (on average) one synaptic vesicle—often none, sometimes two or three; most synapses are not terribly reliable. One synaptic vesicle contains about 5,000 molecules of a neurotransmitter such as glutamate, and this is released into a narrow synaptic cleft, acts on receptors on the postsynaptic site, and is rapidly removed by transporters to be recycled. Everything is over in a few milliseconds. In the synaptic cleft, the concentration of neurotransmitter reaches very high levels, and the receptors at which it acts require these high concentrations. Peptide vesicles carry a much larger cargo (about 85,000 molecules), and their receptors are sensitive to concentrations a thousandfold lower than receptors for neurotransmitters. Peptides are broken down slowly, with half-lives that are generally a few minutes—at least 10,000 times longer than those of neurotransmitters.
Moreover, peptides are not necessarily released at synapses in the first place. As in the excerpt above, molecules released at synapses into the synaptic cleft are pretty likely to wind up at one particular target neuron. But neuropeptides can be released from any part of a neuron—not just synapses but also cell body and dendrites. In the latter cases, once released, the molecules just wander off in any direction. (You might have learned in high school that the dendrites are the inputs of a neuron, not the outputs. Ha! Not for neuropeptides.)
So the upshot is: peptides are well-suited for broadcasting signals, but wholly inappropriate for doing the heavy-duty computation (probably trillions of calculations per second) involved in sensory processing, motor control, search, planning, and so on. The latter is the domain of the classic neurotransmitters—glutamate, GABA, acetylcholine, etc.
2.4 …Yet one pool of neurons can still send three independent signals simultaneously using the same neuropeptide
In Chapter 19, Leng discusses the connection between oxytocin and eating in rats. The story there winds up being pretty simple. When a rat is hungry, it’s a good time to seek food, and a bad time to seek friendship and sex. When a rat is full, it’s the other way around. Thus, the big (“magnocellular”) oxytocin neurons that I mentioned above have receptors for α-MSH, a neuropeptide released in the brain after eating. Under most conditions (but not pregancy), the α-MSH triggers the release of oxytocin into the brain, which then in turn triggers behavior that promotes friendship and sex. Makes sense!
(This story is specific to rats. In humans, eating leads to the secretion of vasopressin not oxytocin.)
Anyway, this brings him to an interesting point: in rats, the same group of magnocellular oxytocin neurons is controlling three processes simultaneously using just that one neuropeptide:
Releasing oxytocin into the brain, where it helps control behavior;
Releasing oxytocin into the body, where it controls certain digestion-related processes like sodium excretion (by the kidneys into the urine, a.k.a. natriuresis) and gastric motility;
Releasing oxytocin into the body, where it controls the “milk letdown” reflex in the mammary glands of lactating females, or uterine contractions during birth.
How does that multiplexing work? Leng has a good answer:
The neurons release oxytocin into the body from axons during spikes, and the same neurons release oxytocin into the brain from dendrites through a process unrelated to spiking.
The neurons release oxytocin into the body in the form of a constant baseline plus periodic pulses. The kidney ignores the pulses but responds to the constant baseline, while the mammary glands (for lactating females) or uterus (for pregnant females) ignores the constant baseline and responds to the pulses.
By the way, related to this last part, Leng spends maybe 10% of the book walking us through the decades-long journey to figure out the nuts-and-bolts low-level mechanics of exactly how a heterogeneous, loosely-connected, noisy group of oxytocin neurons could emit coordinated bursts every few minutes, creating the oxytocin pulses that trigger milk let-down. (If you want to skip to the conclusion, here’s his 2008 computational model.) I don’t know why he and everyone worked so hard on that. Who cares exactly how the oxytocin neurons synchronize their bursts? It seems to be an idiosyncratic “implementation detail” that doesn’t matter for anything else. So here we have yet another excellent example of why building brain-inspired Artificial General Intelligence would be infinitely easier than understanding the brain.
3. Case study of hypothalamus “business logic”: NPY/AgRP neurons
(Note: This section also draws from the paper “Understanding how discrete populations of hypothalamic neurons orchestrate complicated behavioral states”, Graebner et al. 2015.)
Within the arcuate nucleus of the hypothalamus is a subpopulation of neurons that produce the neuropeptide NPY (“neuropeptide Y”, where I think “Y” is somehow related to its 3D shape), and AgRP (“agouti gene-related peptide”, where agouti is a type of fur coloration, don’t ask me why), and GABA (“γ-aminobutyric acid”, which is not a neuropeptide but rather a “conventional” inhibitory neurotransmitter).
When you stimulate these NPY/AgRP neurons, the animal eats more. So these neurons are unusually well-studied, thanks to their presumed relevance to obesity.
Here is a (very incomplete) list of properties of these neurons, and how they correspond to legible, evolutionarily-plausible, “business logic”:
On the input side:
Ghrelin is emitted by an empty stomach into the bloodstream. When ghrelin binds to NPY/AgRP neurons, it makes them express more mRNA for NPY & AgRP, and also makes them fire more. Or more simply, the “business logic” pseudocode is:
“When your stomach is empty, other things equal, you should eat more.”
Leptin is emitted by fat cells into the bloodstream. When leptin binds to NPY/AgRP neurons, it makes them express less mRNA for NPY & AgRP, and also makes them fire less. Or more simply:
“When you have lots of fat cells, other things equal, you should eat less.”
Peptide YY is a hormone emitted by the gut after you eat. Like leptin, it inhibits NPY/AgRP neurons.
“When you just ate, other things equal, you should eat less.”
On the output side:
NPY/AgRP neurons send axons to the parabrachial nucleus (PB) of the brainstem. To a first approximation,[3] PB tracks homeostatic state variables, to be used in homeostatic feedback control. One lab found that upon destroying AgRP neurons, mice would starve to death, but if they injected imitation-GABA into the corresponding part of PB, the mice would eat again. Therefore, presumably, the AgRP neurons are telling PB (via GABA) that the current homeostatic state is “undernourished”. Or more simply:
“When you’re undernourished, relay that fact to the brainstem’s homeostatic state-estimation systems.”
PB is also tracking whether the current homeostatic state involves signs of injury (and if so, it triggers various signals related to “pain”). The same[4] NPY/AgRP axons in PB are releasing not only GABA (as above) but also NPY, and it turns out (ref) that some injury-related neurons in PB have receptors for NPY that change their activity, with the result of suppressing pain-related behavior. Or more simply:
“When you’re undernourished, increase pain tolerance—getting food is more urgent than long-term investment in health by tending to injuries.”
NPY/AgRP neurons send axons to the periaqueductal gray (PAG) in the brainstem. Not coincidentally, some PAG neurons have NPY receptors. What does PAG do? To a first approximation, if PB is the brainstem center in charge of detecting homeostatic problems, then PAG is the brainstem center in charge of correcting them;[3] it’s a major brainstem motor output center, particularly (but not exclusively) involving autonomic nervous system activity. I can’t find great information about what the NPY-sensitive PAG neurons do, but this paper speculates that maybe they’re controlling sympathetic nervous system processes that are especially energy-consuming, like nonshivering thermogenesis. OK sure, that seems as good a guess as any, although I suppose there are other possibilities too. More simply:
“When you’re undernourished, reduce homeostatic activities that consume a lot of energy.”
NPY/AgRP neurons project to a few places in the telencephalon—the ones I know about are the central nucleus of the amygdala (CeA), the BNST, and the lateral and medial septum (source 1, source 2). I have thoughts about what these signals do and how, but it’s a long story and I need to nail down the details and check it more carefully. I’ll defer that discussion to another post on another day. Sorry!
NPY/AgRP neurons inhibit α-MSH neurons. α-MSH generally has the opposite effects to NPY & AgRP, i.e. it reduces feeding, and causes various other changes appropriate to being full (e.g. lower stress, higher sex drive), via lots of specific connections and receptors paralleling those above.
“When you’re undernourished, turn off the internal signal that says that you’re not undernourished!”
I don’t know all the mechanisms—and some may be indirect—but “previous studies demonstrate that central injection of AgRP neuropeptides inhibit puberty and sex hormones, reduce nociception, increase locomotion, and increase plasma stress hormone concentrations.” (source)
“More generally, when you’re undernourished, downweight long-term investments in health and reproduction, and instead prioritize uses of energy that further the cause of immediate food-seeking.”
Conclusion: Why did I go through all that? Because I wanted to give you a better feel for what the hypothalamus does and how.
To be sure, that’s an incomplete accounting of the functions of one little cell group among many dozens (or even hundreds?) in the hypothalamus. So yes, these things are complicated! But they’re not hopelessly complicated. Keep in mind, after all, the entire brain and body needs to get built by a mere 25,000 genes. My current low-confidence feeling is that reasonably-comprehensive pseudocode for the human hypothalamus would be maybe a few thousand lines long. Certainly not millions.
4. More potential “gotchas” for would-be hypothalamus scholars
Conveniently, many neuropeptides have descriptive names. For example, CRH stands for “Corticotropin-Releasing Hormone”, and you guessed it, it’s a hormone that triggers the release of corticotropin. But not-so-conveniently, Leng has a throwaway comment when talking about prolactin-releasing peptide that “as is often the case with the names of neuropeptides, this name is inappropriate”. :-P
It may be unwise to think too hard about why a neuron might be expressing a small amount of some peptide or receptor. It could just be random and functionally-irrelevant, he argues:
“It might be relevant that all magnocellular vasopressin cells also express some oxytocin, though mostly very little, while all magnocellular oxytocin cells express some vasopressin, though again mostly very little. It’s hard to believe that this coexpression is functionally meaningful. … [Similarly,] all of the α-MSH cells appear to make some AgRP and NPY, but usually at very low levels, while those that make large amounts of NPY and AgRP all appear to produce some small amounts of α-MSH. … Perhaps the cost of evolving ways of repressing such expression completely is not worth the modest cost of some promiscuous but biologically irrelevant expression. Not everything produced by a cell necessarily matters; gene expression is, like everything in a cell, noisy and messy.”
5. Conclusion
The book was a bit scattered and disorganized, and had some rambly digressions thrown in that left me saying “what on earth is that doing there?”. But to be fair, I could say those same things about the hypothalamus itself.
Anyway, I found the book a lovely visit into a fascinating field of research, by an expert who has spent a long career in the trenches and who I am strongly inclined to trust.
I was left feeling that really understanding how and why the hypothalamus does something-in-particular—starting from individual protein interactions and cascading all up to behaviors—is very hard, but possible, or at least it’s possible when enough people care to spend years or decades working on it. Quite a bit less hard is to understand generally what particular neurons are doing and why it’s evolutionarily useful, without piecing together all the gory details of low-level mechanisms.
I’ve commented before that, to my dismay, the most theory-minded, AI-adjacent neuroscientists are especially likely to spend their talents on what I call the Learning Subsystem (neocortex, hippocampus, cerebellum, etc.), while almost entirely ignoring the hypothalamus & brainstem, except of course where they interface most directly with the Learning Subsystem (e.g. the brainstem’s dopamine neurons that transmit reinforcement-learning-related signals).
I can understand where they’re coming from. Stare at the Learning Subsystem for long enough, and you find beautiful algorithms with wide applicability—algorithms that in some cases may be transferable straight into future revolutionary AI algorithms. By contrast, stare at the hypothalamus for a decade, and you can learn one hyper-specific fact about how oxytocin neurons coordinate to fire in synchronous bursts that lead to milk let-down. Or you can unravel the gory molecular details of how hunger increases pain tolerance, or whatever.

Nevertheless, I argue here that the “business logic” part of the brain, and probably the hypothalamus in particular, is hiding some fascinating algorithm-level secrets yet to be revealed which will be highly relevant for safe and beneficial AI—secrets related to the symbol grounding problem, reward function design for reinforcement learning, the core logic underlying human social and moral intuitions, and so on. So I strongly encourage all those neuroscientists with a knack for algorithms and AI to not forget about the poor hypothalamus. Maybe start by reading this book!
Needless to say, there’s much, much more in the book that didn’t make it into the review above. For example:
Prairie voles, when they first meet, have sex more or less continuously for about thirty-six hours.
We still have much to learn.
(Thanks Linda Linsefors & Thang Duong for critical comments on a draft.)

- ^
My sense right now is that the “business logic” in the brainstem is a bit more tilted towards innate lower-level sensory-processing and motor programs—like which muscles to contract when vomiting, or a multi-step calculation that guesses the presence/absence of a slithering snake in the field-of-view—whereas the “business logic” in the hypothalamus is kinda more like implementing a high-level controller. We’ll see an example in Section 3. See also Larry Swanson’s “Cerebral hemisphere regulation of motivated behavior” (2000) for a similar picture in which “hypothalamic controllers” sit on top of the brainstem motor system hierarchy.
- ^
The closest thing I could find was Neuron Numbers in the Hypothalamus of the Normal Aging Rhesus Monkey, which estimates 6,000,000 neurons in the rhesus monkey hypothalamus, including a helpful partial breakdown by nucleus.
- ^
I’m getting this mainly from the book How Do You Feel by Bud Craig.
- ^
Well, probably it’s the same neurons, but it’s also possible that there are multiple subpopulations of NPY/AgRP neurons doing different things within PB. I didn’t check.
- 8 examples informing my pessimism on uploading without reverse engineering by (3 Nov 2023 20:03 UTC; 126 points)
- Connectomics seems great from an AI x-risk perspective by (30 Apr 2023 14:38 UTC; 107 points)
- [Intro to brain-like-AGI safety] 3. Two subsystems: Learning & Steering by (9 Feb 2022 13:09 UTC; 106 points)
- Spatial attention as a “tell” for empathetic simulation? by (26 Apr 2024 15:10 UTC; 55 points)
- My AGI safety research—2022 review, ’23 plans by (14 Dec 2022 15:15 UTC; 51 points)
- I’m confused about innate smell neuroanatomy by (28 Nov 2023 20:49 UTC; 40 points)
- Connectomics seems great from an AI x-risk perspective by (EA Forum; 30 Apr 2023 14:38 UTC; 10 points)
- 's comment on AXRP Episode 22 - Shard Theory with Quintin Pope by (21 Jun 2023 17:35 UTC; 6 points)
- 's comment on Is RL involved in sensory processing? by (30 Oct 2022 3:55 UTC; 4 points)
- 's comment on Biomimetic alignment: Alignment between animal genes and animal brains as a model for alignment between humans and AI systems. by (EA Forum; 30 May 2023 18:24 UTC; 2 points)
That confirms what I have been getting around to over time: Human instincts and motivational systems are probably built from few elements. I used to think that there are hard-wired motivations/interests for specific types of sports or hobbies or people and how this could be coded for. But after reading Steven’s sequence, I look more for simple patterns like geometric features, hormonal triggers, or sensory triggers that could—after a lot of learning—give rise to such preferences. Maybe some sport is only interesting because it involves fast-moving objects in the visual field and the smell of grass…
This study has a table that lists the half-lives of many neuropeptides (see table 2):
https://febs.onlinelibrary.wiley.com/doi/full/10.1111/j.1742-4658.2011.08051.x
It would be interesting to compare the half-lives to the duration of behaviors, emotions, and moods associated with these neuropeptides. I only recognize the NPY mentioned above in the study.
I remember being fascinated with the potential to help fix problems for people, especially eating problems, by adjusting the levels of various neuropeptides. I still think neuroscience ought to boldly tamper with those for the sake of making peoples’ lives less miserable, but now I’m much more fascinated by the implications for setting up a control system for RL-based AI. I also wonder if anyone has already made something like a simplified-business-logic model of the functions of the hypothalamus.